



Håller homologiargumentet?


I oktober 2005 hölls en debatt i tidningen Dagen om evolutionsteorins förhållande till kristen tro. Jag vill i denna artikel mycket kort fokusera på ett argument som återkommande ansågs bevisa darwinismen som teori, nämligen det så kallade homologiargumentet. Argumentet säger i korthet att likhet innebär släktskap.
När man jämför likheter mellan organ hos olika arter kan lätt vissa problem uppstå, eftersom likheterna kan vara svåra att definiera, och det är ofta mycket svårt att kvantifiera skillnaderna mellan organismerna. Istället studerar man numera DNA-sekvenser, eftersom man där kan göra omfattande statistiska beräkningar och relativt lätt ställa upp s.k. fylogenetiska evolutionsträd. Låt oss titta närmare på homologiargumentet, eftersom det ofta anses bevisa att människor och apor är nära släkt med varandra. Argumentet ser ut som följer:
- Organismer vars DNA liknar varandra delar en gemensam förfader. Ju mer lika organismerna är, desto mer nyligen har denna förfader levt.
- Apor (i synnerhet schimpanser) och människor är genetiskt lika varandra.
- Därför delar apor och människor en gemensam biologisk förfader.
Argumentet kan verka självklart och övertygande, men det finns anledning att vara skeptisk till slutsatsen.
Den första premissen
Den första premissen, att organismer som liknar varandra delar en gemensam förfader, kan t ex ifrågasättas. Inom fylogenetiken (den del av molekylärbiologin som försöker ställa upp evolutionära träd, huvudsakligen baserat på DNA-sekvenser) skiljer man mellan homologi och homoplasi. Homologi är när en egenskap hos två organismer liknar varandra, därför att de båda har ärvt den från en gemensam förfader. Homoplasi är när organismerna uppvisar likhet som inte beror på gemensamt ursprung.
Klassiskt skiljer man mellan tre olika fall av homoplasi: parallell evolution (organismerna har förvärvat egenskapen från samma egenskap hos förfäderna), konvergent evolution (organismerna har förvärvat egenskapen från olika egenskaper hos förfäderna) samt sekundär förlust (en tillbakagång till ett tidigare stadium i utvecklingslinjen, vilket gör att organismen kan börja uppvisa samma egenskap som andra organismer).
Intelligent design-teorin och skapelseteorin har en diametralt annorlunda uppfattning och menar att det finns ett femte sätt att förklara genetiska likheter på som varken omfattas av homologi eller homoplasi: en medveten design.
Den första premissen - att organismer som liknar varandra genetiskt delar en gemensam förfader - är alltså inte empiriskt sann eftersom det finns en alternativ tolkning (se figur 2). För att få den empiriskt sann, kan man vända på premissen och säga att organismer som delar en gemensam förfader liknar i allmänhet varandra genetiskt (med några lysande undantag så vitt man vet)1. Då blir förvisso premissen empiriskt sann, men istället följer inte slutsatsen av premissen, eftersom det även finns andra sätt att förklara likhet på (t ex homoplasi och design) och argumentet faller likväl.
Påståendet att alla hästar är däggdjur är sant, men inte det omvända - alla däggdjur är inte hästar. På samma sätt bör organismer som delar en sentida gemensam förfader likna varandra, men inte nödvändigtvis tvärtom - att två organismer liknar varandra innebär inte med automatik att de delar en gemensam förfader (homologi) eller ens att de har kommit till genom evolution (homoplasi). Det är t ex möjligt att en designer på ett eller annat sätt har orsakat människors och schimpansers genetiska skillnader. Detta kan naturligtvis rent logiskt ha skett genom evolution, men en designer är inte bunden till att skapa genom evolution.
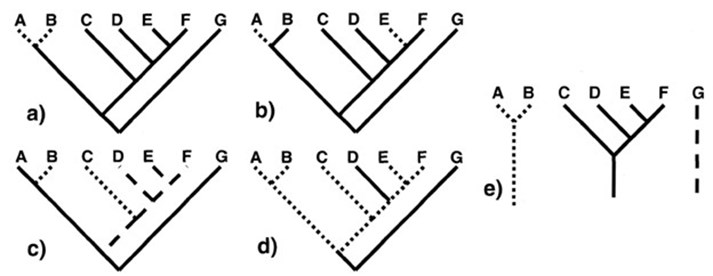
a.) I det här exemplet skiljer sig A och B från gruppens gemensamma förfader och sägs därför vara apomorfiska. Egenskaperna C, D, E, F och G är däremot primitiva och kallas för plesiomorfa.
b.) Separat utveckling (homoplasi) från samma egenskap hos A och E kallas för parallell evolution.
c.) Separat utveckling (homoplasi) från olika egenskaper hos B och C kallas för konvergent evolution.
d.) D uppvisar en sekundär förlust och återgång till en tidigare egenskap i utvecklingskedjan och får därmed samma egenskap som G (homoplasi).
e.) Enligt intelligent designteorin och skapelseteorin kan likheter förklaras genom en designers medvetna val. Dessa likheter kan vara oberoende av evolution. När det handlar om små förändringar (mikroevolution) ger homologi sannolikt en korrekt bild av organismernas utveckling i de flesta fall, men för större förändringar (makroevolution) kan man misstänka att likheterna beror på att designern har orsaker som är oberoende av evolution.
Om det finns tillräckliga skäl för en designer att använda sig av likheter, och dessa skäl är oberoende av evolutionära mekanismer, vill jag hävda att detta är tillräckligt för att motivera en icke-evolutionär tolkning av genetiska likheter. Därmed är inte heller den första premissen giltig och slutsatsen i argumentet faller. Finns det då några sådana skäl? Svaret är ja, och detta kan exemplifieras med varför mänskliga designers använder sig av likheter i olika produkter. Detta kan t ex vara för att kunna utnyttja effektiva standardkomponenter i olika sammanhang, göra dem enhetligare, elegantare och lättare att förstå. Det är inte alls omöjligt att en designer av samma skäl skulle använda sig av likheter i sina skapelser, oberoende av tillverkningsmetoden (evolution eller inte).
En viss likhet är också nödvändig om världen ska kunna fungera (tänk om alla individer hade levt på olika sockerarter och aminosyror) och ofta kan likheter mellan olika produkter vara ett sätt för tillverkaren att marknadsföra sig, som kung Salomo skrev:
"Ur storheten och skönheten hos det skapade träder bilden av dess upphovsman fram för tanken."
Med tanke på att det finns miljoner arter i världen, är det dessutom ofrånkomligt att vissa arter är mer lika varandra än andra. Självklart kommer det att vara så!2
Alltså, även om organismer som delar en gemensam förfader liknar varandra, finns det andra skäl, och goda sådana, till varför de skulle kunna göra det utan gemensamt biologiskt ursprung som bakomliggande orsak. När de darwinistiska mekanismerna (mutationer och naturlig selektion) å andra sidan är tillräckliga för att förklara organismernas tillkomst, finns det inget vetenskapligt tvång att tillgripa en designer, men självklart kan en designer ha verkat ändå, eftersom en designer alltid kan imitera naturliga processer, och som vi precis har sett finns det mycket goda skäl för en sådan att använda sig av likheter och dessa skäl har ingenting med evolution att göra.
Homologi och homoplasi är alltså enbart giltiga när vi har system som darwinismen klarar av att åstadkomma. I övriga fall, t ex när genprodukterna uppvisar specificerad komplexitet3, ger homologi eller homoplasi sannolikt ingen korrekt bild av deras tillkomst. När man inte kan använda darwinistiska mekanismer för att förklara komplexa biologiska system, bör man alltså inte heller använda homologi eller homoplasi för att försöka förklara organismernas släktskap.
Ovanstående resonemang kan tyckas självklart, eftersom homologi förutsätter ett evolutionärt släktskap, men poängen är att även om man argumenterar att en designer har styrt makroevolutionen och man på så vis kan förklara uppkomsten av specificerat komplexa molekylära system som en darwinistisk evolution aldrig skulle kunna förklara, bör homologiargumentet ändå inte användas som stöd för detta. Varför inte? Därför att det finns goda skäl för en sådan designer att använda sig av likheter, och dessa skäl är inte på något sätt kopplade till evolutionen.
Homologi och homoplasi kan alltså ersättas av en alternativ tolkning av genetisk likhet som ett medvetet val av en designer. Den första premissen i homologiargumentet är därmed ogiltig.
Den andra premissen
Ovanstående är egentligen tillräckligt för att visa att slutsatsen inte gäller, men låt oss ändå titta lite närmare även på den andra premissen. Att apor (i synnerhet schimpanser) och människor är genetiskt lika varandra är ganska säkert. Frågan är hur lika. Nyligen har arvsmassan hos schimpanser kartlagts och de spännande resultaten presenterades för några veckor sedan i Nature4. Som kristna kan vi självklart vara mycket entusiastiska åt detta - den första uppgiften som Gud gav människan var ju just att studera och klassificera djuren och evolutionära forskare har nu fullgjort denna befallning åtminstone för schimpansen bättre än någonsin tidigare i världshistorien. Det handlar dock än så länge bara om en mycket osäker sekvens, en "draft sequence" som sannolikt innehåller en hel del fel och kan komma att ändras framöver. De preliminära siffrorna visar dock att skillnaden är större än vad som har sagts tidigare. När man vanligtvis .hör siffran 1,23 % (som döljer drygt 40 miljoner kvävebaser!) är enbart substitutioner inräknade, men här är även andra skillnader, t ex repeterade sekvenser, insertioner, deletioner, omorganiserade kromosomer m.m. också inkluderade.5 En del av detta är svårt att kvantifiera, men rapporter har kommit från evolutionskritiskt håll som menar att det rör sig om åtminstone 7 % skillnad.6

Den stora diskussionen bland forskare idag är dock hur stor betydelse DNA:t egentligen har för individens egenskaper. Troligtvis är det inte lika mycket som man tidigare har trott, med tanke på hur stora skillnader det är mellan apor och människor på anatomisk, psykologisk, kulturell, andlig m fl. nivåer. Det är framförallt på dessa högre organisatoriska nivåer som man får söka svaret på vad som gör människan unik. Hälften av allt DNA hos människan är t ex identiskt med det hos bananen, men det innebär inte att människan är hälften banan. På samma sätt innebär inte DNA-likheterna mellan människa och schimpans nödvändigtvis att människan är 93 % schimpans.7 Varför betraktas då människan som så nära släkt med schimpansen? Antropomorfismen inom primatforskningen baserar sig knappast på objektiv vetenskap, utan på ett nödgat paradigm, där design som förklaring till genetiska likheter utesluts i metodologisk reduktionistisk anda. Detta kan naturligtvis inte tas som ett argument mot skapelseberättelsens historicitet. Som Stanley Sjöberg uttryckte det i debatten i Dagen (21/10), är kritiken mot Bibeln av filosofisk karaktär och inte vetenskaplig.
Slutsatsen
Slutsatsen i homologiargumentet, att apor och människor delar en gemensam förfader, är inte giltig eftersom premisserna inte stämmer med verkligheten. En annan minst lika giltig tolkning är att de delar en gemensam designer och en sådan inte är beroende av att skapa genom evolution. Tvärtom bör vi förvänta oss att en designer använder sig av likheter av andra orsaker. Denna tolkning är särskilt trolig om slumpmässiga evolutionära processer inte kan åstadkomma den skillnad som finns mellan människor och apor, vilket allt fler forskare menar är fallet. I ett sådant läge är en designer nödvändig och en sådan behöver inte skapa genom evolution.
Fakta talar aldrig för sig själva utan tolkas alltid i ett större sammanhang, ett paradigm. När evolutionsträd byggs upp, oavsett om det baseras på homologi eller homoplasi, är det baserat på förutsättningen (som är just en förutsättning och inte en slutsats) att evolutionen har ägt rum men för att få svar på om det verkligen är så, måste man gå utanför detta paradigm. Jag har här inte argumenterat för att denna förutsättning är felaktig, men homologiargumentet bevisar den inte och kan aldrig göra det heller, eftersom evolutionen är en premiss och inte en slutsats. Då får man leta någon annanstans.
Noter
- Se Ibstedt, S. Vad säger DNA-likheterna om människan och hennes ursprung? Genesis 2003(4):12-20.
- Se vidare Ibstedt, S. ibid.
- Se Ibstedt, S. The Design Revolution. Genesis 2005(2):18-21.
- The Chimpanzee Sequencing and Analysis Consortium. Initial sequence of the chimpanzee genome and comparison with the human genome. Nature 2005;437:69-87. doi:10.1038/nature04072.
- Se även Ibstedt, S. 2003, ibid och DeWitt, D.A. Människa-apa likhet > 98 % - inte nu längre. Genesis 2005(3) s. 26. Dessa artiklar är dock skrivna innan hela genomet var färdigsekvenserat.
- Chimpanzee Genome: Evidence for Evolution? Reasons to Believe; nyhetsbrev 051024. http://www.reasons.org/resources/new_reasons
- Ibstedt, S. 2003, ibid.
 Tidigare publicerad i: Genesis 2005/4
Tidigare publicerad i: Genesis 2005/4
Denna artikel är tidigare publicerat i magasinet Genesis 2005/4.
PrenumereraFler nummerRelaterat Innehåll
 Homologi på lätt sätt
Homologi på lätt sätt
När evolutionister hittar anatomiska eller biokemiska likheter mellan organismer som de menar har gemensamt ursprung, så presenteras likheterna som om de bevisade detta släktskap. Generna eller organe ...
Artikel | 10 Apr 2020
 Är inte likheterna mellan människa och apa vad gäller anatomi, socialt betende m.m. bevis för evolution?
Är inte likheterna mellan människa och apa vad gäller anatomi, socialt betende m.m. bevis för evolution?
Det finns likheter mellan människa och apa vad gäller anatomi, socialt betende (t.ex. språk, moral etc.). Är inte denna likhet bevis för en evolution? ...
FAQ-sida | 6 Apr 2013
Genesis Nyhetsbrev
Kommande händelser
- 6 May 2024, 00:00: Göran Schmidt har temadagar för studenter på YWAM Transform, Stockholm
- 17 Oct 2024, 12:00: Nordic Creation Research Conference 2024: "A creationist perspective on the origin of diversity in biological life"
- 18 Oct 2024, 18:00: Genesis Årskonferens / Skapelsekonferens 2024


Föredrag av Stuart Burgess, professor i mekanisk design vid Bristoluniversitetet, på Skapelsekonferens ...

Tolvte delen i en föreläsningsserie om Biblisk Kreationism. Producent: Ulf Fagerberg - ulf@kristentv.s ...

Del 3 i en videoserie i fem delar med Anders Gärdeborn, med namn "24 myter om evolutionen".A Se hela s ...
Frågor & Svar
- Vad säger ni om Ron Wyatts påståenden om att man hittat Noas Ark?
- Är universum slutet eller öppet?
- Hur bestämmer man ålder på berg? Finns det några bevis på att bergen inte är så gamla som man tror inom evolutionsteorin?
- Hur kan Bibeln säga att regnbågen blev till först efter Noas flod?
- Var "Lucy" mänsklig?
