



ERV


Endogena Retrovirus (ERV) – argumentet för gemensamt ursprung som istället blev argument för gemensam designer.
Du som har följt med inom genetikområdet har kanske hört att endogena retrovirus (ERV) har fungerat som ett vanligt argument för den evolutionära idén om gemensamt ursprung, i synnerhet mellan människa och schimpans, de senaste åren. ERV:er, som utgör omkring 8% av vårt DNA, sägs vara rester av virusangrepp i förhistorisk tid, som har integrerats i vår arvsmassa på exakt samma platser som hos andra djur, men i varierande grad och mest likt just schimpansen. Det anses därför bevisa att vi har ett nära släktskap. Även mutationer i samma delar av ERV ger stöd för tolkni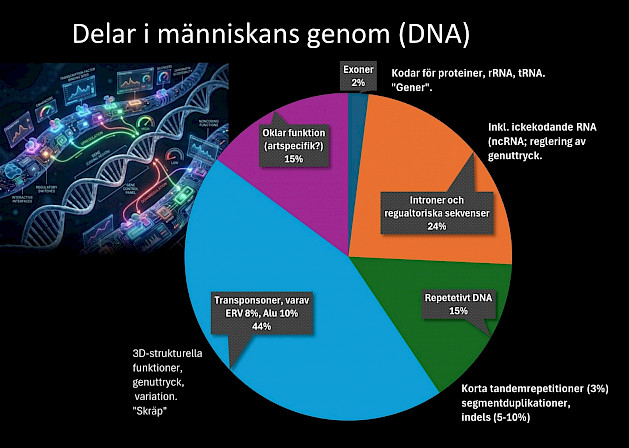 ngen.
ngen.
Med en evolutionär syn är detta ett ”vattentätt” argument för gemensam härkomst mellan människor och aporna. Men forskningen på de delar av DNA som inte kodar för proteiner, det som allmänt ansågs vara evolutionära rester, och brukade benämnas ”skräp-DNA”, har gått rasande snabbt framåt. Idén om skräp i vår arvsmassa har hämmat forskningen med decennier, så ännu är det mycket vi inte vet, men en sak står klar. Det finns inget skräp. Forskarna hittar nya funktioner hela tiden i det man tidigare förmodade var virusrester, så själva namnet ”endogena retrovirus” får många att le lite överseende över den totala bristen på insikt det vittnar om. Ändå upprepas påståendena om gemensamt ursprung med hänvisning till ERV:er så ofta, att ett klargörande är nödvändigt.
Bilden intill visar översiktligt vad vi idag vet om innehållet i vårt DNA. I varje cell finns ungefär 2 meter DNA som totalt har 3 miljarder baspar i dubbel uppsättning, tätt ihoprullat på 46 kromosomer (23 par) där omkring 20 000 proteinkodande gener finns. Av dem är flera procent unika för människan. Sekvenser som liknar virus utgör ca 8%, eller 4 gånger större andel av DNA än generna.
Om Endogena RetroVirus (ERV) som evolutionärt argument - i ett nötskal
ERV:er är funktionella, designade DNA‑element. Det vi vet om deras funktion stödjer inte gemensam härkomst, utan separat ursprung och designad biologisk mångfald. Den evolutionära tolkningen bygger på föråldrade antaganden om ”skräp-DNA”.
Likheter hos olika organismer eller i genetiken används ofta som evolutionärt argument, men är exakt vad en modell som bygger på skapelse förväntar sig också. Inom design finns alltid likheter och hierarkiska mönster så argumentet svarar inte på om evolution eller skapelse är orsaken. I stället är funktionell genetik den avgörande diskriminerande faktorn mellan evolution och skapelse, inte likhet eller hierarkiska mönster.
Argumentet för ett evolutionärt ursprung utifrån ERV:er
Argumentet som används för gemensamt ursprung är att dessa DNA-sekvenser är rester av forntida retrovirusinfektioner som infogats i könsceller och ärvts vertikalt. Delade ERV‑positioner mellan olika arter skulle innebära att de fått infektioner på samma plats i DNA, vilket är osannolikt om inte arterna har gemensamt ursprung. Man studerar mutationer i ERV/LTR‑sekvenser och använder dem som molekylära klockor för att se när arternas grenar skildes från varandra.
Tre argument stödjer påståendet om att ERV beror på gemensam härkomst:
- Samma ERV på samma genomposition
- Likartade mutationer i ERV‑sekvensen
- Hierarkisk fördelning av dessa mutationer
Den grundläggande invändningen är att alla dessa argument bygger på förutsättningen att ERV:er är icke‑funktionella skräprester, vilket är falskt.
Designad variation - epigenetik
Utifrån ny kunskap om epigenitik vet vi nu att stor biologisk variation kan ske otroligt snabbt, utan någon förändring av den genetiska koden i DNA. Det är tydligt att när Gud skapade de olika organismerna fanns förprogrammerad genetisk variationsmöjlighet som kan uttryckas både genom olika epigenetiska faktorer och olika alleler (genvarianter). Skapelsemodellen förutsäger att hög heterozygositet (många genvarianter) fanns redan från början, vilket ger variationsmöjligheter. Biologisk variation kräver inte lång tid och mutationer. Snarare visar studier på motsatsen.
Observerad biologisk variation är förväntad i en skapelsemodell och är inte bevis för ett gemensamt ursprung.
Argument som inte ger stöd åt något håll, men ofta används
Likhet (homologi) och hierarkier kan förklaras både av evolution och design. Dessa är därför icke‑diskriminerande.
En analogi är hur fordon (bil, SUV, traktor) uppvisar likhet p.g.a. funktion, inte släktskap. Vetenskapens system för klassificering (t.ex. däggdjur) är konstruerade av människan och ger därför inte i sig själva bevis på härkomst, även om människan där klassas som apa.
Det finns likheter, men vi kan inte bortse från mycket stora skillnader mellan människan och schimpansen.
Förutom 10-12% skillnad i längd hos DNA och åtminstone 400 miljoner ”bokstävers” (nukleotiders) skillnad, vet vi också att det är ett kraftigt avvikande genuttryck i hjärnan, olika alternativ splitsning och stora skillnader i reglerande DNA mellan arterna. Det epigenetiska mönstret stämmer dåligt med evolutionära förväntningar[i] och människans 3D‑genomstruktur är unik.
Likhet kan alltså lika gärna indikera gemensam design som gemensamt ursprung.
Vederläggning av ERV‑argumentet
Om ERV:er är funktionella och nödvändiga då kan de inte vara evolutionära skräprester.
Den vetenskapliga litteraturen visar att
- ERV:er reglerar genuttryck
- Är avgörande för embryonal utveckling (t.ex. placenta, syncytin)
- Bidrar till immunsystemet (antiviral effekt)
- Delar av ERV (solo‑LTR) är aktiva regulatoriska element
- ERV:er formar kromatinarkitektur och celltypdifferentiering
ENCODE‑projektet har hittills visat att ~80 % av genomet visar biokemisk aktivitet. Hela skräp‑DNA‑paradigmet är kraftigt ifrågasatt.
Evolutionära försök att rädda sin teori går ut på att virus ”återanvänds”, dvs virusresterna har ”utvecklat funktion”. Detta är en efterhandskonstruktion som saknar experimentell demonstration och bara fungerar som ett retoriskt nödlösningsargument.
Mutationsargumentet (LTR‑hierarkier)
Det evolutionära påståendet är att delade mutationer i ERV/LTR visar på gemensamt ursprung.
Men modern forskning visar att mutationer inte är slumpmässiga, utan att det finns mutationshotspots. En mycket uppmärksammad studie visade att funktionellt viktiga gener har lägre mutationsfrekvens, och att DNA-reparationssystemet prioriterar vissa regioner framför andra[ii]. Epigenetik påverkar var mutationer sker och många mutationer kan vara designade variationer.
Hierarkiska mönster kan uppstå utan gemensamt ursprung
Alu‑sekvenser och andra transposoner
Tidigare har stora delar av DNA betraktats som parasitiskt skräp, bland annat Alu (hoppande gener), men studier har visat att de reglerar RNA‑bearbetning, påverkar alternativ splitsning, är viktiga för stressrespons, har antivirala effekter och är mer aktiva i människa än schimpans.
Delade Alu‑positioner är därmed ett starkt argument för gemensam design, inte gemensamt ursprung.
Var kommer virus från?
Retrovirus kan ha uppstått från genomets egna flyttbara element. När cellen flyttar dessa element bildas alla komponenter som ingår i virus. Vid en virusinfektion tar virusets arvsmassa över kontrollen av en cell och får den att producera nya virus, så funktionen finns redan i cellen. Virus är i så fall degenererade avknoppningar av flyttbara element. I så fall fanns ERV:er före virus, inte tvärtom [iii]Detta vänder helt på evolutionens orsak–verkan‑modell.
Sammanfattande slutsats
- ERV:er är nödvändiga funktionella komponenter
- Skräp‑DNA‑paradigmet är vetenskapligt föråldrat
- Gemensamt ursprung kräver stora mängder funktionslöst DNA som inte finns.
- När funktion påvisas → kollapsar argumentet för common descent
Gemensamma DNA‑element är därför ett starkt argument för gemensam design. Inte gemensam härstamning.
Artikeln bygger på The Endogenous Retrovirus Handbook av Donny Budinsky.
[i] Helgenomstudier av DNA‑metylering i hjärnan visar att hundratals till tusentals gener har signifikant olika metyleringsmönster mellan människa och schimpans. Skillnaderna är särskilt koncentrerade till promotorer och regulatoriska regioner, inte slumpmässigt spridda. Gener med epigenetiska skillnader är kraftigt anrikade för neurologiska, kognitiva och psykiatriska funktioner. (Zeng et al. 2012 – helgenom‑metyleringskartor av mänsklig och schimpanshjärna, https://pmc.ncbi.nlm.nih.gov/articles/PMC3511995/ ; Martin et al. 2011 – stabila, artspecifika epigenetiska tillstånd i människoapor, https://pubmed.ncbi.nlm.nih.gov/21908772)
[ii] I Arabidopsis thaliana uppstår mutationer preferentiellt i mindre viktiga genomregioner, medan essentiella gener skyddas genom effektivare DNA‑reparation.
CpG‑öar muterar 10–50 x oftare än andra regioner p.g.a. metylering och spontan deaminering.
Repetitiva element (LINE, Alu, LTR) är tydliga hotspots.
[iii] Borger, P. (2025). Reassessing Viral Origins: From Escaped Genes to Degenerated Microbes. Pathogens, 14(12), 1205. https://www.mdpi.com/2076-0817/14/12/1205
 Magnus Lindborg
Magnus LindborgOrdförande för Genesis.
Relaterat Innehåll
 Transposonens dag 16:e juni
Transposonens dag 16:e juni
Idag, 16:e juni är det transposonens dag, något som uppmärksammas bl.a. av tidskriften Mobile DNA, för att hedra minnet av Barbara McClintock, som upptäckte dessa fascinerande "flyttbara" DNA-sekvense ...
Nyhetsartikel | 16 Jun 2025
 Likhet mellan människa och schimpans kraftigt överdriven visar ny forskning
Likhet mellan människa och schimpans kraftigt överdriven visar ny forskning
Uppgifterna om 98% sekvenslikhet mellan människa och schimpans är direkt felaktiga, särskilt när de anges utan sin kontext. Man har valt att göra jämförelserna på ett sätt som gör att intrycket av den ...
Artikel | 22 Mar 2022
Genesis Nyhetsbrev
Kommande händelser
- 23 Oct 2026, 18:00: Skapelsekonferens / Genesis Årskonferens 2026


Föredrag med Anders Gärdeborn på Skapelsekonferens 2018 i Östersund. (Observera att Anders inte förme ...

Josef frågar Göran om genetik och de ökande problem som finns för den som vill mena att människan utve ...

0:00:00 Inledning. Vem är du, Rebecca? 0:03:50 Varför är du inte darwinist? 0:05:05 Bevisen som gick ...
Frågor & Svar
- Har allt land suttit ihop i "superkontinenten" Pangea?
- I böcker kan man hitta nästan kompletta utvecklingen för hästen. Vad säger ni om det?
- Hur kunde människan leva tillsammans med en dinosaurie som Tyrannosarus Rex (T-Rex)?
- Det sägs att människans och schimpansens DNA 99% likt - är inte det ett bevis för att de är släkt?
- Finns det hemliga koder i Bibeln?
